
The just concluded SICB meeting in Salt Lake City featured 19 talks on anoles. You can see the titles and authors here, or go to here to check out the abstracts. Anyone who attended want to tell us about the meetings?
Category: New Research Page 66 of 67

Anolis takes its rightful place on the cover of PNAS. Photo of A. distichus vinosus by R. Glor.
Larger islands have more species. Why? MacArthur and Wilson’s theory explains island species richness as an equilibrium between the input of new species (a function of island isolation) and extinction (inversely related to island area). Although certainly one of the most influential ideas in biology in the 20th century, the theory had its limitations, most specifically, that it relied solely on ecological phenomena—colonization and extinction—to explain species richness. Yet, that can’t be the whole story, because islands are renowned for their evolutionary exuberance—witness the adaptive flowering of lemurs on Madagascar, finches in the Galápagos, honeycreepers on Hawaii and so on. MacArthur and Wilson were, of course, well aware of the evolutionary component of island diversity and discussed the need to incorporate evolutionary issues into their theory at the end of their monograph.

Anolis grahami and A. lineatopus
Research on the escape behavior of lizards has become somewhat of a cottage industry in the last two decades, with scores, if not hundreds, of papers examining the effect of factors such as temperature, concealment, and crypticity. Probably the most important early paper in this area (and perhaps the first period) was Stan Rand’s study of the effect of body temperature on flight initiation distance of Anolis lineatopus. This work—conducted on the grounds of the University of the West Indies in Mona (a suburb of Kingston), Jamaica—reported that lizards with lower body temperatures fled at greater distances from an approaching predator. Rand speculated that this pattern resulted because warmer lizards could run faster, setting the stage for the pioneering work on the effect of temperature on sprint locomotion by Ray Huey, Al Bennett, and others.
More than four decades later, Bill Cooper returned to the scene of Rand’s work to further study the escape behavior of A. lineatopus and its relative A. grahami. Following the method used by Rand and many since, Cooper walked directly toward lizards at a constant pace and noted how far away he was when they fled, as well as the manner in which they escaped. Although the two species differ in habitat use, A. grahami being more arboreal, escape behavior was very similar. In both species, lizards tended to escape by running up trees, often by moving to the far side of the tree (termed “squirreling” by many anole aficionados); lizards initially perched lower in the vegetation tended to initiate escape at greater distances; and lizards in areas with greater human activity appeared to be habituated to the presence of people and delayed escape until the faux predator was relatively close.
None of these results is surprising; rather, they agree quite closely with work on other anoles and other types of lizards. Cooper makes an interesting observation that anoles that flee to the ground, such as grass-bush anoles, show an opposite pattern, fleeing at greater distances when they are perched higher in the vegetation. This, of course, makes sense because the higher they are, the further they are from safety, the opposite of the relationship that occurs in species that flee upward. As Cooper notes, more comparative work on other species, both more types of ecomorphs and species from other islands, could prove instructive. In addition, studies using non-human predators would also be welcome to establish the extent to which behavior elicited in response to approaching humans is representative of how anoles respond to their natural predators. Other studies have used snake or bird models to study anole escape behavior. In this paper, Cooper explains why he and others use humans for these tests—ease and repeatability of methods are certainly major advantages. Nonetheless, research on other types of predators would be an interesting avenue for future work.
Finally, Anole Annals awards a booby prize to the copy editor of this journal for the unique distinction of having a typo in the first line of the abstract (“fight” instead of “flight”) and what appears to be a sentence fragment that was supposed to have been deleted as the first words of the article itself.
{kind=link}
 Darwin’s finches are the iconic example of adaptive radiation. Some researchers, including me, have had the temerity to suggest that the diversification of Caribbean anoles may join the finches as an exemplar case study. But just how similar are these two radiations, in terms of evolutionary pattern and process? And can we learn anything from a two-clade comparison? I explore these questions in a chapter in a recently released book that resulted from a symposium held two years ago to honor Peter and Rosemary Grant. My chapter concludes:
Darwin’s finches are the iconic example of adaptive radiation. Some researchers, including me, have had the temerity to suggest that the diversification of Caribbean anoles may join the finches as an exemplar case study. But just how similar are these two radiations, in terms of evolutionary pattern and process? And can we learn anything from a two-clade comparison? I explore these questions in a chapter in a recently released book that resulted from a symposium held two years ago to honor Peter and Rosemary Grant. My chapter concludes:
“Overall, adaptive radiation in Darwin’s finches and Greater Antillean anoles has occurred in very much the same way. Interspecific competition appears to have been the driving force leading to resource partitioning and subsequently adaptation to different niches, and speciation is probably primarily allopatric and may be promoted as an incidental consequence of adaptation to different environments. Differences exist as well, such as the extent of hybridization and of independent evolution on different islands; many of these differences probably result because the radiations differ in age and aspects of natural history.”
If you want to read the whole thing, it’s available here.
Incidentally, the book, In Search of the Causes of Evolution: From Field Observations to Mechanisms, presents a nice overview of the breadth of evolutionary biology, with chapters by workers as diverse as Dolph Schluter, Andy Knoll, Cliff Tabin, David Jablonski, Scott Edwards, David Wake and Hopi Hoekstra, among others.

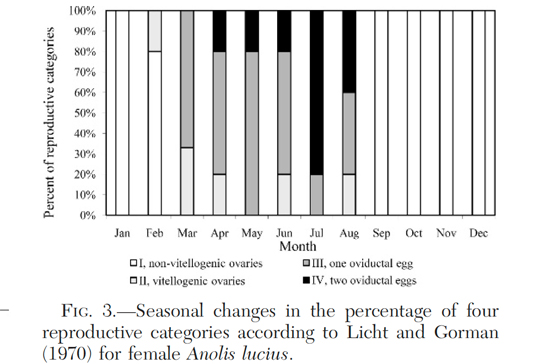
Many lizard species lay one or two multi-egg clutches each year; anoles, however, distribute their egg laying over the course of several months by producing a single egg every week or two. Although this unusual aspect of anole reproduction is conserved across the entire genus, other aspects of anole reproduction exhibit considerable variation. The annual reproductive cycle of anoles, for example, is known to vary from nearly continuous year-round egg production to highly seasonal reproduction limited to the warmest or wettest months. This variation appears to result from a combination of regional environmental variation, reproductive cycle plasticity, and historical contingency. In the latest issue of Herpetologica, Domínguez et al. (2010) provide new details on the reproductive cycle of female A. lucius. Although some previous reports have suggested continuous reproduction in Anolis lucius, Domínguez et al. find that female reproduction in populations near Havana, Cuba is highly seasonal; all specimens examined had non-vitellogenic ovaries between September and January before reaching peak egg production in July. Although their quiescent period is shorter, the reproductive cycle of A. lucius is similar to that of the better-studied temperate species A. carolinensis in being driven by photoperiod and temperature. Two other noteworthy facts seemed worth sharing. First, like other anoles from the northern Neotropics, male and female A. lucius reach maturity in approximately eight months. Second, communal egg-laying in A. lucius is noteworthy because, like other rock-dwelling species from Cuba (i.e., A. bartschi and A. argenteolus), females often lay in small cavities in cliffs, caves and rocks rather than in soil or trees.
{kind=link}
Imagine you’re walking down a busy city street, talking with your friend about the latest post you read on Anole Annals. You’re getting to the most exciting part of the discussion when a bus rumbles by and drowns out your sentences. Your friend can’t hear you and loses out on amazing anole knowledge.
Animals that live in complicated environments with noisy backgrounds face similar communication challenges. The signal can be auditory – imagine a frog trying to find a mate in a multispecies chorus in a Costa Rican swamp that just received its first rain of the year. The signal can be olfactory – imagine an ant trying to follow an hour-old trail laid down by one of its sisters. The noise can be visual too – imagine a transparent jellyfish trying to find a mate in the poor light environment of the ocean.
Or, Waldo trying to find Wenda.

Tough, right?
To ensure that communication is successful, animals should evolve signals that are highly conspicuous in the environment in which those signals are used.
Anoles are well known for their charismatic displays. They do push-ups, bob their heads, and extend a colorful flap of skin beneath their chin called a dewlap. Anoles display during territorial conflicts, courtship behavior, and predator avoidance. Anoles tend to live in very noisy environments. For example, how is a male anole displaying in the low-light forest understory to be seen against a background of shaking leaves?
One way for an anole to be seen is to create a dewlap whose color has great contrast in the light environment in which it is usually found. Another way is to use a display with jerky movements and rapid dewlap extensions that stands out even against low light and shaking leaves (Ord and Stamps 2008). A third way to be seen, discussed in a new paper by Ord et al. (2010), is to time displays to run during periods in which background noise is relatively low (i.e. the wind dies down and the leaves stop shaking). Imagine pausing your city-street conversation of anoles until after the bus had passed.
Ord et al. test whether anoles match their displays to calm backgrounds by comparing the display timing of Jamaican anoles, which use a rapid, jerky display, to Puerto Rican anoles, which use a slow, drawn out display. They hypothesize that the Puerto Rican species, with their slower head bobs and dewlap extensions, will time their displays to match a quiet background. In contrast, Jamaican species, whose display is thought to be suitable in noisy visual conditions, should show less background matching (Ord and Stamps 2008).
To answer this question, Ord et al. analyzed videos of lizard displays, comparing background noise during lizard displays to similar stretches of time when lizards were not displaying. They compared four species on Jamaica to four species on Puerto Rico. The species were similar in habitat use and dewlap color between islands, and there were no appreciable differences in the physical environment between islands.
Consistent with their hypothesis, Ord et al. found that Puerto Rican species actively avoided displaying during periods of high environmental noise; they waited until until the visual background calmed down. In contrast, Jamaican lizards did not time their displays as carefully. Thus, it appears that these two lineages of Anolis lizards have evolved different strategies to solve the problem of how to communicate in noisy visual environments.

Puerto Rican anoles avoid displaying during periods of high noise.

The distichus group on Hispaniola, including forms suggested for full species status by Glor and Laport. Image from their Figure 1.
In a recently accepted paper in Molecular Phylogenetic and Evolution, Rich Glor and Robert Laport suggest that the stalwart Hispaniolan anoles, A. distichus and A. brevirostris, actually represent complexes of species under the general lineage species concept. This idea is not new, but it’s surprising how long it’s been since it’s gotten serious attention. Before his tragic death in a car accident, Preston Webster made substantial progress working on the brevirostris-group species problem, using allozyme data to study the genetics of Haitian populations in the 1960s and 70s. Webster found evidence for genetic isolation among several brevirostris populations differing in dewlap color (Webster and Burns 1973), but never erected any species. He advocated the idea though, and under the guidance of legendary Caribbean systematist Albert Schwartz, Douglas Arnold erected three additional species from within Anolis brevirostris, naming one after the late Preston Webster (A caudalis, A. marron, and A. websteri; Arnold 1980). He also named erected two additional A. brevirostris subspecies.
In several later papers, Susan Case also used allozymes to study distichus and brevirostris subspecies, but she stopped short of naming any as species. She did, however, note evidence of stable genetic breaks among some distichus subspecies (e.g., Case and Williams 1984).
In the paper at hand, Glor and Laport turned their attentions mainly to the widespread Hispaniolan Anolis distichus, which has 16 subspecies (including some not on the main island of Hispaniola) that often differ remarkably in dewlap coloration. Focusing on the Dominican Republic side of Hispaniola, they found evidence for deep divergence in mtDNA among these subspecies, and argue that they represent distinct evolutionary lineages. The picture isn’t completely clean of course – one of the forms (favillarum) renders another (dominicensis) paraphyletic for this particular marker – but the authors still make a case that these groups represent true evolutionarily distinct units. Certainly some will disagree with Glor and Laport’s decision to elevate these subspecies using the general lineage concept. But species concepts aside, it’s hard to argue with their conclusion that “studies that continue to treat A. distichus (sensu lato) as a single reproductively continuous unit will be problematic.”
Check out the paper! I think this is the tip of the iceberg for distichoid anole systematics. Glor and Laport refrained from tackling the Haitian distichus subspecies and also remained agnostic on the formal status of the three subspecies of A. brevirostris. Rest assured this is just the beginning though…

The green thing on the left doesn’t look like a real anole to me. But apparently it’s convincing enough to both predators and other anoles, because both attack them, as evident by the bite marks they leave on these clay doppellizards. McMillan and Irschick deployed these faux lizards in trees in two areas in New Orleans, on the Tulane campus and in a wilder area outside of town. In the wild area, the models had bites marks clearly identifiable as belonging to lizard jaws (photo on right), and others that were from birds or other presumed predators. By contrast, perhaps not surprisingly, on the lovely grounds of Tulane, models were bitten by anoles, but not by predators. Also not surprisingly, there were no bites of any kind in either locality in the winter, and many more in the spring and fall. You might say to yourself: maybe lizards and predators just like to bite clay or anything new that shows up in their environment. How do we know that they actually thought they were attacking a lizard? Well, we don’t, of course, but the researchers did put out cubes made of the same material, and none of these were attacked, which lends some credence to the possibility that these models, cartoonish as they seem, may have passed for the real thing. McMillan and Irschick, following others, suggest that these models may serve as a proxy for levels of predation and intraspecific aggression.
Anoles only lay one egg at a time. This penurious habit has been speculated to result from their arboreal lifestyle—because they spend so much time running around on vertical surfaces, they can’t afford to be weighed down by a big clutch of eggs. Reasonable enough, but is there any evidence that carrying eggs has any cost to female anoles? In a recent study in Ethology, Johnson et al. find that in A. carolinensis, as the mass of the egg increased, display behavior and movement rates decreased, suggesting that females are less active as they become more gravid. Whether this is inactivity results because females are more vulnerable to predators or for other reasons is not known.

Photo of A. schwartzi copyright J. Losos.
This post is for those of you who might not be regular readers of Amphibia-Reptilia. Last summer, Joyce et al. reported on habitat mediated variation in size, population density and mating system in the St. Eustatius (Lesser Antilles) anole A. schwartzi. Joyce et al. quantified body size, density, and frequency of display behaviors in populations of A. schwartzi occupying habitats with and without available rocky substrates. They found denser populations of larger individuals that tend to display more frequently occupying rocky substrates. Their results suggest that the mating system and intensity of competition for mates varies among habitats in a predictable manner. This article is another in a long string of valuable contributions from Bob Powell’s REU courses.