
Hello Anole enthusiasts. This will be a slightly different post to the usual in that Anolis won’t feature at all! I am one of those weird lizard researchers that is yet to feel the scientific attraction many of my colleagues feel towards Anolis and, as such, have always preferred their much spikier old world agamid counterparts- the garden lizards of the genus Calotes.
I’m currently doing a Ph.D. (supervised by long term AA member Adam C. Algar at The University of Nottingham, UK) that looks at how agamids use the various habitat types within South East Asia (in particular, Peninsular Malaysia), focussing on oil palm plantations, rubber plantations and secondary forests. While some agamids occupy one or more of these habitat types, only a single genus has colonised all of them- Calotes. Calotes versicolor will doubtless be familiar to any herper who has been on vacation to Southern Asia, a large, red-headed take on Anolis sagrei with the same aggressive character and, unfortunately for everything else, the same ability to dominate in invasive situations. While C. versicolor is present in human modified habitats in Malaysia, the forested North of the country is home to a larger, even spikier species: C. emma, which will occupy the forest edges that C. versicolor seems to avoid. We chose C. emma for this reason as the ability to draw comparisons between the semi-natural forest edges and man-made plantations was key to this research.

An adult female Calotes emma in a heavily disturbed oil palm plantation (photo: James J. Hicks)
We measured such Anolis study staples as body temperature (Tb), perch type and behaviour when encountered and all the usual morphology traits. We also characterised habitats structurally using random transects and thermally using ibuttons in copper models and measured thermal performance using a racetrack (not an easy piece of equipment to build in the tropics!) and HD camera.
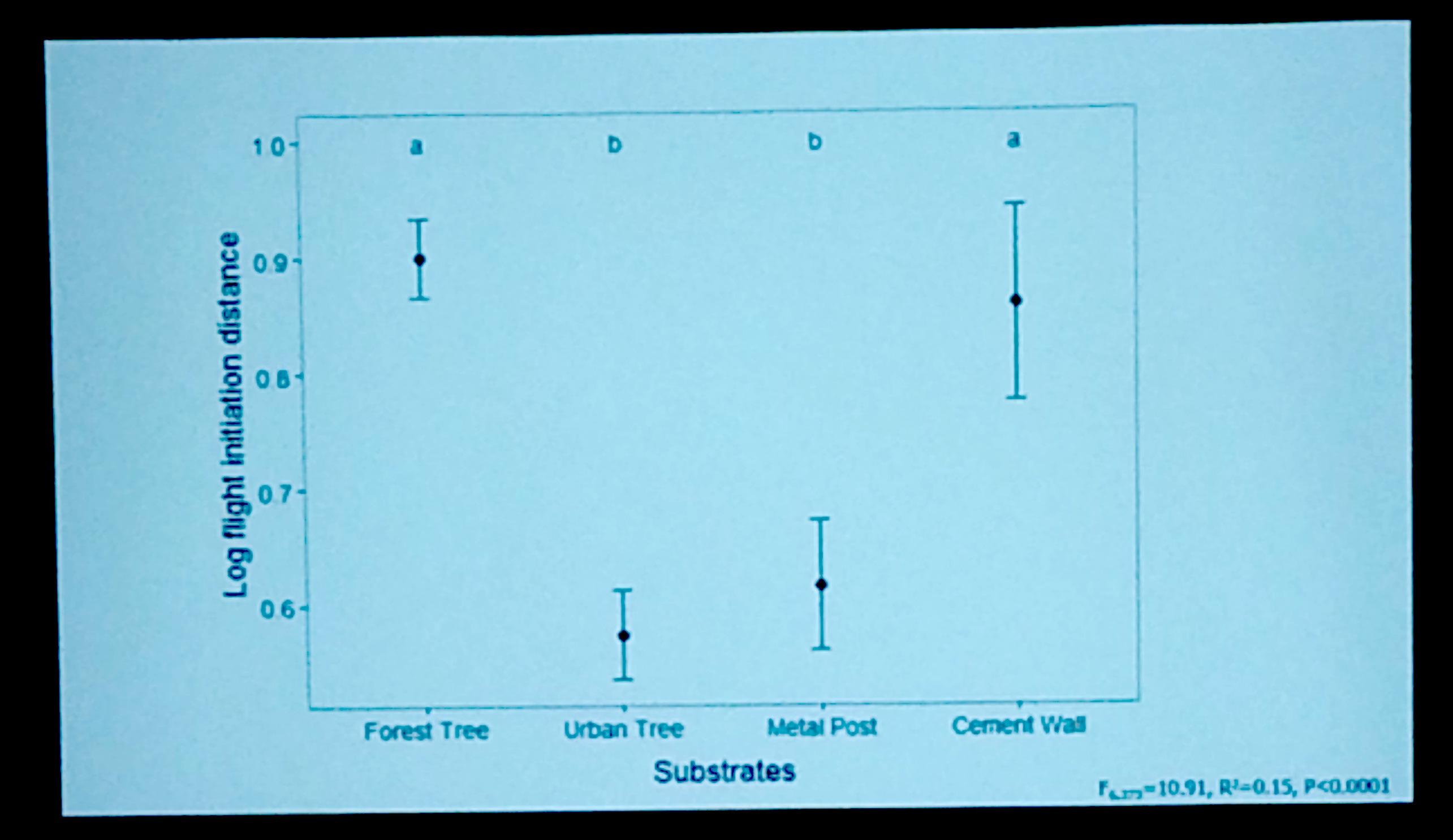
Differences in behaviour were noticed immediately with C. emma being highly arboreal in rubber plantations (and difficult to noose!), using the smooth trunks to ascend from predators/herpers and rotating around in typical agamid fashion. In oil palm plantations the equivalent ‘trunk’ structures consisted of blocky remnants of fronds that form a hard, smooth surface that lizards tended to avoid (shame these ones don’t have toepads!). Consequently most C. emma were seen on the ground and in and amongst discarded piles of the fronds at most ca. 40 cm from the ground, using these piles as both perches and refugia. In forest edge habitat Calotes used a wide range of perches that incorporated similar heights and structures to those used in the plantation habitat but was much less abundant than in either plantation type. These behavioural differences coincided with differing femur lengths with the arboreal Calotes in rubber having the longest femurs, for their snout-vent length alongside utilising statistically wider perches.
Thermally, plantation habitats were hotter and more variable than forest edges at relevant scales to lizards as has been documented previously at larger scales. Despite this, operative temperatures fell well within C. emma’s thermal tolerances and, currently at least, seem more favourable for this species. Whether climate change will push these habitats closer to C. emma’s critical thermal maximum remain to be seen but applying the ‘standard’ 3°C rise still won’t, theoretically at least, impact their thermal performance.
In short, plantations seem to be great habitat for Calotes emma! They are extremely abundant in these man made habitats, more so than any other vertebrate species despite each plantation type seemingly forcing them to rely on different, single axes of their fundamental niche. The downside is, as always, plantations trade low abundance and high diversity for high abundance and low diversity. While rubber plantations support at least 3 other agamid species in our study area, C. emma was the sole representative in oil palm. Whether this is due to thermal aspects (is it too hot for larger-bodied forest dragons?) or structural aspects (Draco probably can’t glide onto oil palm trunks) or something completely different remain to be tested in a future session of fieldwork. This will focus on assessing the more poorly understood forest specialist agamid species’ structural and thermal niche axes and whether these requirements are met by plantation habitats.

My future fieldwork will focus on forest specialist agamids such as Gonocephalus bellii (pictured) and try to explain why they are absent from human dominated habitat types (photo: James J. Hicks)

























{kind=link}