

Male Anolis nebulosus
With few exceptions, there has been far less research on the ecology and evolution of anoles inhabiting islands off Central and South America compared to anoles on islands in the West Indies. Siliceo-Cantero, Garcia, Reynolds, Pacheco and Lister (2016) have recently taken a step towards leveling the playing field by making a detailed comparison of a mainland A. nebulosus population residing in the Chamela-Cuixmala Biosphere Reserve in western Mexico, and a population inhabiting the small island of San Agustin in the Bay of Chamela just 5 km from the mainland study site. Their research focused on three central questions: (1) To what extent has the island population diverged morphologically, behaviorally, and genetically from the ancestral mainland population? (2) Does the morphology of the island and mainland populations correspond to any of the six West Indian ecomorphs?, and (3) In the absence of mainland competitors, has the island population undergone niche expansion and evolved towards a generalist phenotype?
Initial analyses of morphology indicated that both island and mainland populations were dimorphic, with each sex in each location having a large-headed, short-limbed morph, and a less common small-headed, long-limbed morph. The only other anole known to exhibit such intra-sexual dimorphism is Anolis agassizi from Mapelo Island, where there are large and small male morphs that also differ in coloration. In keeping with a pattern often found in anoles, island and mainland A. nebulosus morphs showed a significant positive correlation between average size-adjusted upper limb lengths and the mean logarithm of their perch diameters, suggesting that the morphs may be adapted to different microhabitats. Island morphs were also significantly larger than their mainland counterparts, and both the CV and Shannon diversity of the SVL distribution of all four morphs combined were significantly greater for the island population. The authors calculated the area of morphospace occupied by the morphs, as defined by the first two axes of a principal components analysis based on several morphological characters. On both island and mainland, the existence of a second morph doubled the total morphological space of each population. With respect to divergence in shape, mainland male and female morphs had significantly longer snouts and wider mandibles than the corresponding Island morphs. A PCA clearly separated island and mainland males from males of all the West Indian ecomorphs.

Female A. nebulosus
The genetic divergence of the island and mainland populations was assessed by sequencing both mitochondrial and nuclear genes. Genetic diversity of mtDNA was low on the island (haplotype diversity (h)= 0.50; nucleotide diversity (π) = 0.0007) relative to the mainland (h= 0.95; π= 0.006), and the populations were fairly divergent (Fixation Index = 0.34). Haplotype diversity of nuclear DNA, however, was somewhat higher on the island (h = 0.38 vs. .32). Based on 16,382 SNPs, island and mainland individuals were grouped cleanly by both Discriminant Analyses of Principal Components and STRUCTURE, a Bayesian clustering algorithm, and had 100% posterior support in STRUCTURE for membership in their respective populations. The authors conclude that these results indicate incomplete lineage sorting in mtDNA, as well as a relaxation population genetics model in which the island population is evolving independently of the mainland population under the influence of drift and lack of gene flow.
While the A. nebulosus population on San Agustin has undergone niche expansion in the absence of mainland competitors, there was no detectable evolution towards a generalist morphology, defined as a hypothetical phenotype intermediate between the shapes of the six West Indian ecomorphs. The authors suggest, however, that this definition is unrealistic given that the selective forces necessary to produce such a Jack-of-All-Trades ecomorph are most likely absent on the small islands harboring solitary Anolis populations. Alternatively, they propose that reduction in death rates, higher densities, increased intraspecific competition, and greater male-male competition for females are the major selective forces common to many small islands. A large body of theory and data indicate that all of these forces can drive the evolution of larger body sizes and greater SSD. The authors point out that Schoener’s work documenting wide-spread convergence in body size and SSD among solitary anoles in the West Indies, supports this hypothesis, and predict that the distributions of body size and SSD on single species continental islands should converge on those found in the West Indies (see Figure below).
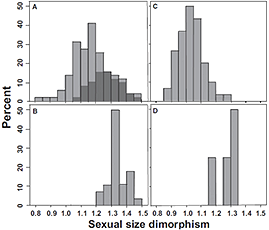
A. Distribution of SSD indices for 102 Anolis species from the Greater Antilles. Dark grey bars delineate trunk-ground and trunk-crown ecomorphs. B. SSD distribution for 28 solitary West Indian anoles (data from Schoener, 1977). C. SSDs for 60 mainland species. D. Histogram of SSD for four solitary species including A. nebulosus, inhabiting islands off Central and South America (data for A. concolor and A. pinchoti from Calderon-Espinosa and Barragan-Forero, 2011. Data for A. agassizi from Lopez-Victoria et al., 2011). After Fig. 3 in Siliceo-Cantero et al, 2016.
 Three years ago I received a message from Ray Huey asking me if I’d be interested in collaborating on an NSF grant to return to Puerto Rico and replicate the studies on Anolis thermal biology and ecology that we had conducted during the 1970s. The idea was to have the original investigators, including Ray, myself, and Paul Hertz, work in the same study areas, utilize the same techniques, and document changes that had occurred over the past 35-40 years. Our ultimate goal was to understand the impact of climate warming on Anolis populations over a range of habitats, from the Luquillo rainforest in northeastern Puerto Rico to the Guanica dry forest in the southwestern corner of the island.
Three years ago I received a message from Ray Huey asking me if I’d be interested in collaborating on an NSF grant to return to Puerto Rico and replicate the studies on Anolis thermal biology and ecology that we had conducted during the 1970s. The idea was to have the original investigators, including Ray, myself, and Paul Hertz, work in the same study areas, utilize the same techniques, and document changes that had occurred over the past 35-40 years. Our ultimate goal was to understand the impact of climate warming on Anolis populations over a range of habitats, from the Luquillo rainforest in northeastern Puerto Rico to the Guanica dry forest in the southwestern corner of the island.