

Lineage accumulation curves (Fig. 2, Rabosky and Glor 2010) showing that Hispaniola (blue), Jamaica (purple), and Puerto Rico (orange) have reached speciation-extinction equilibrium. Cuba (red) is still gaining species.
Losos, J.B., and D. Schluter. 2000. Analysis of an evolutionary species-area relationship. Nature 408: 847-850.
{kind=link}
Losos and Schluter (2000) return to Caribbean anoles to test three hypotheses about the species-area relationship: (1) that there is an area threshold above which speciation surpasses immigration as a source of new species; (2) above the threshold size, speciation events per unit time should increase with island area; and (3) the slope of the species-area relationship should become steeper above the area threshold. Qualitatively similar to Losos (1996), this paper was novel in that a newly available, nearly complete, mt-DNA phylogeny allowed Losos and Schluter to reconstruct immigration and speciation events and to model whether species number has reached speciation-extinction equilibrium.
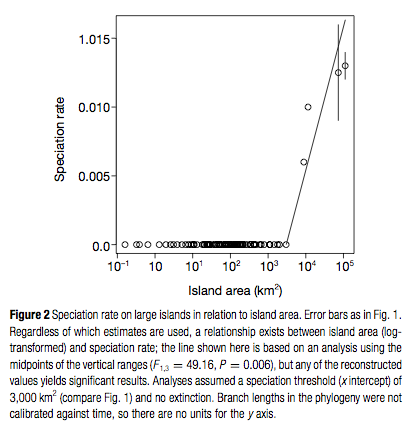
The authors found that speciation is the dominant source of new species for the four islands of the Greater Antilles. For example, the authors infer that the ten species on Puerto Rico (8,959 km2) arose from three ancestral lineages that experienced seven in situ diversification events. On Cuba (114,525 km2), 42 of the 57 species belong to two of the clades that have radiated on Cuba, while 12 other species also likely resulted from in situ speciation. In contrast, of the 143 islands smaller than 3000 km2, no in situ diversification was inferred, again suggesting a minimum island size below which speciation does not occur for anoles. The authors also found an increase in speciation rate with island area (hypothesis 2) and that there was a significant difference in the species-area relationship between Greater Antillean islands and the small islands (hypothesis 3). Hence, the authors provide more evidence that an evolutionary perspective needs to be included in any consideration of the species-area relationship.
 A key point in any estimation of diversification rates and speciation events from molecular phylogenies regards extinction. Extinction events do not appear on molecular phylogenies so the number of reconstructed speciation and immigration events is likely underestimated. The authors assume, however, that extinction doesn’t act differentially for lineages that appeared due to immigration compared to those that arose in situ, so the ratio of speciation to immigration events plotted against area should still correctly determine the area threshold. As more evidence that extinction is unlikely to be an issue in their analyses, the authors fit a constant birth-death process model to their phylogeny for the Greater Antillean species and find that the best-fit model suggests that speciation increases exponentially through time while extinction is absent. This suggests that the number of Anolis species in the Greater Antilles is still increasing.
A key point in any estimation of diversification rates and speciation events from molecular phylogenies regards extinction. Extinction events do not appear on molecular phylogenies so the number of reconstructed speciation and immigration events is likely underestimated. The authors assume, however, that extinction doesn’t act differentially for lineages that appeared due to immigration compared to those that arose in situ, so the ratio of speciation to immigration events plotted against area should still correctly determine the area threshold. As more evidence that extinction is unlikely to be an issue in their analyses, the authors fit a constant birth-death process model to their phylogeny for the Greater Antillean species and find that the best-fit model suggests that speciation increases exponentially through time while extinction is absent. This suggests that the number of Anolis species in the Greater Antilles is still increasing.
Rabosky and Glor (2010) returned to this last point to re-examine whether diversification is continuing unabated in Anolis or whether the Greater Antillean anoles have reached speciation-extinction equilibrium. Counter to Losos and Schluter, Rabosky and Glor found declining rates of diversification through time and that 3 of the 4 islands have reached equilibrium (see the figure at the top of this post). This paper is detailed in a previous post.
- DNA Sequencing from Formalin-Preserved Specimens - December 16, 2014
- Sans the Claws, Anoles Present Under The Trees? - December 5, 2014
- Out Damned Spot! Or, How They’d Be Loving If They Weren’t Fighting - November 18, 2014
Leave a Reply